植物激素茉莉酸通过核心转录因子MYC2介导的转录重编程调控植物免疫和适应性生长。在静息状态下,抑制子蛋白JAZ直接或通过接头蛋白NINJA 招募转录共抑制子TOPLESS (TPL)来抑制MYC2的转录活性。然而,TPL介导的转录抑制机制以及激素依赖的抑制和去抑制之间的转换机制尚不清楚。
在调控花发育的过程中,TPL招募组蛋白去乙酰基酶(HDAC)对组蛋白进行去乙酰化修饰,来抑制靶基因表达。据此,在现有的茉莉酸信号转导模型中,人们推测TPL采取类似机制招募去乙酰化酶HDA6和HDA19对MYC2靶基因区域的组蛋白进行去乙酰化修饰,从而抑制茉莉酸响应基因的表达。然而有研究表明,HDA6和HDA19的功能丧失突变体均表现出茉莉酸响应基因的诱导表达显著削弱的表型。这与现有模型相矛盾。
中国科学院遗传与发育生物学研究所李传友研究组长期致力于茉莉酸作用机理研究。最近,该研究组重新鉴定了HDA6与HDA19在茉莉酸信号途径中的功能,证实HDA6和HDA19确实是茉莉酸信号途径的正调节因子。研究发现,HDA6和HDA19存在功能冗余,它们对茉莉酸信号的正调控作用并不依赖于其组蛋白去乙酰基酶活性。
进一步的研究发现,HDA6通过与TPL的直接物理互作对TPL进行去乙酰化修饰。HDA6介导的TPL去乙酰化削弱了TPL与接头蛋白NINJA的互作,从而促进TPL从MYC2靶基因处的解离,释放茉莉酸响应基因的表达。同时还发现组蛋白乙酰转移酶GCN5与TPL也存在直接的物理互作,并负责对TPL进行乙酰化修饰。GCN5介导的TPL乙酰化增强了TPL与NINJA的互作,从而促进TPL在MYC2靶基因区域的富集,抑制茉莉酸响应基因的表达。此外,通过质谱等实验鉴定到TPL的关键乙酰化位点K689。该活性位点的乙酰化可促进TPL与NINJA的互作,从而增强对MYC2转录活性的抑制。
有趣的是,HDA6的表达受茉莉酸处理和灰霉菌接种的快速诱导,而GCN5的表达则没有变化。因而,GCN5–TPL–HDA6模块类似于一个分子开关调控TPL乙酰化的稳态,进而灵活地控制茉莉酸信号通路的关闭、开启以及在关闭和开启之间的转换。在静息状态下,GCN5介导的乙酰化作用与HDA6介导的去乙酰化作用使乙酰化的TPL蛋白维持在较高的水平,保持对MYC2的抑制活性,茉莉酸响应基因的表达处于关闭状态。而当植物体内茉莉酸水平响应外界环境刺激而上升时,HDA6的表达被快速诱导而GCN5的表达保持稳定,从而降低TPL的乙酰化水平,削弱其对MYC2的抑制作用,进而促进茉莉酸响应基因的表达(如图所示)。
以上研究是对现有茉莉酸信号转导模型的重要修正和补充,不仅揭示了参与广泛生物学过程的转录共抑制子蛋白TPL对茉莉酸信号通路动态调控的分子机制,还拓宽了人们对乙酰基转移酶和组蛋白去乙酰化酶作用机制的认识。作为动物、植物和微生物中保守的蛋白质翻译后修饰形式,蛋白质的乙酰化修饰参与调控诸多生物学过程。然而相比较动物中非组蛋白的研究,关于植物非组蛋白乙酰化修饰的功能和调控机制还知之甚少,因而该研究为植物中非组蛋白乙酰化修饰的功能和调控机制研究提供了范例。
该研究成果以题为“Regulation of jasmonate signaling by reversible acetylation of TOPLESS in Arabidopsis”于2022年7月3日在线发表于国际著名期刊Molecular Plant杂志 (DOI:10.1016/j.molp.2022.06.014)。李传友研究组安春鹏博士后,邓磊副研究员和博士生翟华伟为论文共同第一作者;李传友研究员为通讯作者。该研究得到国家自然科学基金委和中国科学院战略性先导专项的资助。安春鹏博士得到博士后创新人才支持计划、国家自然科学基金委青年基金和博士后基金的资助。
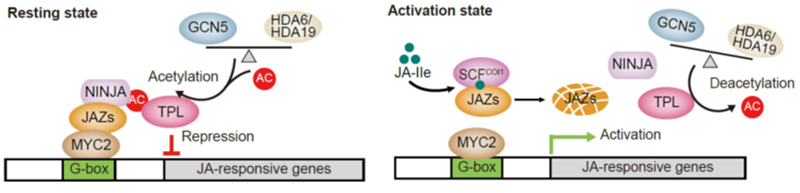
图:GCN5和HDA6介导的TPL乙酰化稳态参与调控茉莉酸信号通路