由于缺乏种子休眠而导致的采前发芽严重威胁着世界范围内的作物生产。作为一种复杂的数量性状,具有适宜种子休眠的作物品种的选育受到有效调控基因的限制。
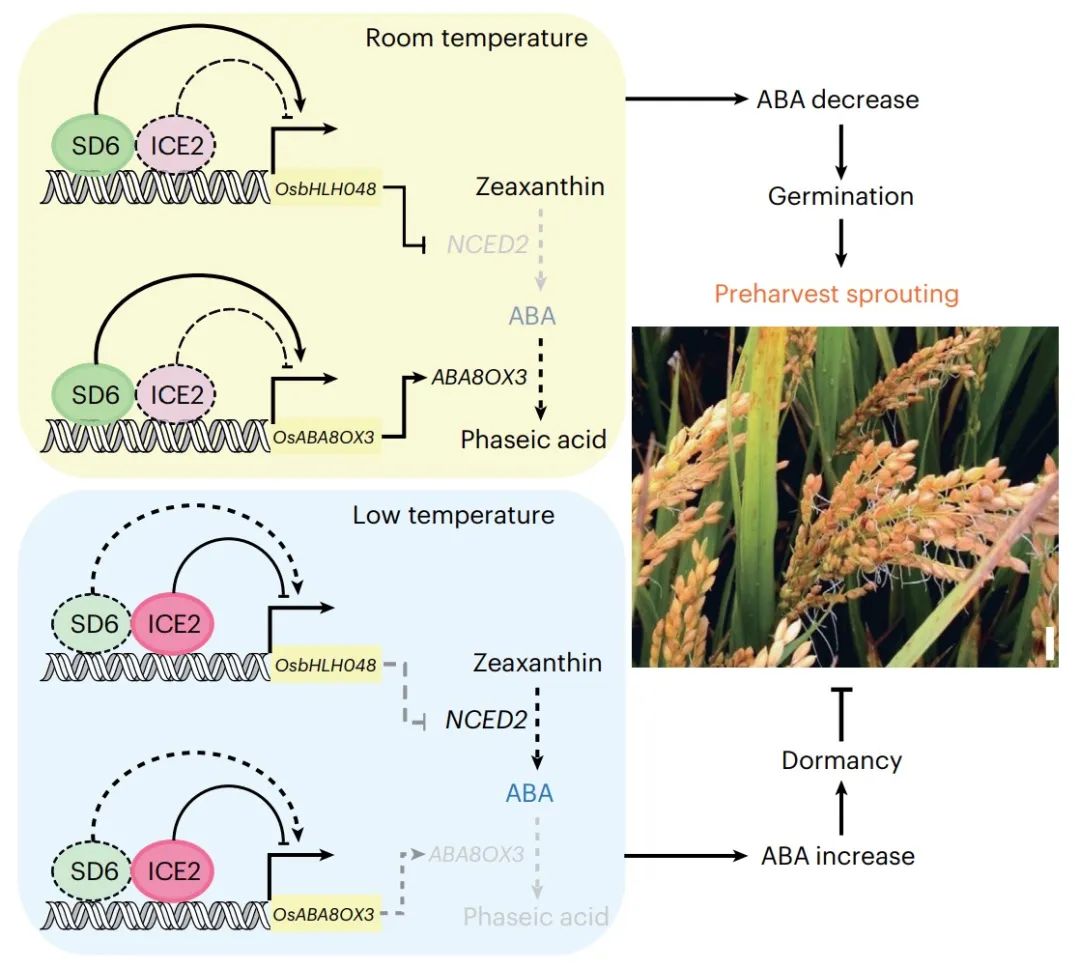
2022年12月5日,中国科学院遗传与发育研究所储成才课题组(现为华南农业大学)与高彩霞课题组合作,在Nature Genetics 杂志在线发表题为“Antagonistic control of seed dormancy in rice by two bHLH transcriptionf actors”的研究论文,该研究成功地将复杂的农艺性状种子休眠分离成一个重要的QTL,种子休眠6 (seed dorcy 6, SD6)。SD6与另一种基本螺旋-环-螺旋(basic helix-loop-helix, bHLH) TF、C-repeat结合因子表达2 (inducer of C-repeat binding factors expression 2, ICE2)诱导子拮抗平衡ABA分解基因ABA8OX3和生物合成基因NCED2的表达,通过响应温度信号控制水稻种子休眠。进一步证实,SD6在水稻和小麦中具有功能保守性,可能是提高田间条件下谷物小灵通抗性的有力靶点。

种子休眠是植物的一种策略,它使种子保持静止状态,直到条件适合发芽。种子休眠影响种子的传播,影响植物生长的位置和时间,因此在自然植物种群中处于强选择状态。充分的休眠使种子在田间处于静止状态,而弱休眠保证播种后的出苗率更高、更均匀。然而,微弱的种子休眠也会导致母体植物上新鲜成熟的种子过早萌发,称为收获前发芽(preharvest sprouting, PHS)。长期以来,包括水稻和小麦在内的谷物小灵通已成为一个严重的世界性问题,导致粮食品质和产量下降。
种子休眠和萌发是由多个数量遗传位点(quantitative genetic loci, QTLs)控制的复杂农艺性状。Gramene QTL数据库记录了164个与种子休眠或萌发相关的水稻QTLs (http://www.gramene.org)。这些QTLs已从栽培水稻、野生水稻和杂草水稻等多种来源报道,但目前在分子水平上鉴定的基因很少。转录因子(transcription factor, TF) Sdr4是受OsVP1调控的种子成熟的全局调控因子。
qSD7-1被追溯至多效基因座Rc,并通过调节ABA生物合成途径控制种子休眠。qSD1-2被鉴定为GA合成基因OsGA20ox2,可能通过GA调节的脱水机制诱导初级休眠。qSd1-1编码与拟南芥DOG1同源的蛋白质OsDOG1L-3,有助于水稻种子休眠的建立。最近,研究人员发现了G基因,该基因通过对ABA生物合成的作用影响种子休眠,并在多个作物科中进行了平行选择。
该研究通过对固定重组个体的可重复表型表征,报道了一个来自aus-type水稻的定量遗传位点,种子休眠6 (Seed Dormancy 6, SD6),编码一个基本的螺旋-环-螺旋(bHLH)转录因子,它是种子休眠自然变异的基础。
进一步研究表明,SD6与C-repeat结合因子表达2 (C-repeat binding factors expression 2, ICE2)的另一种bHLH因子诱导因子(bHLH factor inducer)通过直接调控ABA分解代谢基因ABA8OX3,通过OsbHLH048间接调控ABA生物合成基因NCED2,以温度依赖性的方式拮抗控制种子休眠。SD6弱休眠等位基因在栽培水稻中普遍存在,但在野生水稻中发生负选择。值得注意的是,通过对SD6及其小麦同源物的基因组编辑,证明了SD6是在田间条件下缓解谷物小灵通的有用育种靶点。
总的来说,这项研究结果表明水稻中存在调控种子休眠的SD6/ICE2-OsbHLH048-NCED2级联。与拟南芥同源基因AtbHLH57相比,OsbHLH048在调节水稻种子休眠方面具有相反的功能。因此,这项研究结果揭示了一种新的模式,可以通过不同的机制同时调节植物激素的生物合成和降解,以平衡种子休眠和萌发。
https://www.nature.com/articles/s41588-022-01240-7